…На них надвигалась огромная волна. Однако, всё, что они могли делать – наблюдать, как она стремительно приближается, безжалостно переворачивая привычное равновесие. Они жили вмурованными в двухслойную почву под ногами, оберегали свои врата от внешних посягательств и не впервые переносили подобное цунами, но каждый раз испытывали леденящий всё их естество трепет. В течение лишь нескольких миллисекунд один за одним всё ближе распахивались врата их товарищей, не выдерживая напора, и в них с бешенной силой и победными возгласами устремлялись толпы постоянно дежуривших неподалёку «противников»… Они понимали, что до полной капитуляции оставались считанные мгновения. За их спинами уже расположились налившиеся силой воины, которые были готовы, как только падут последние двери, нестись что есть мочи к следующей крепости, унося с собой необходимые для доставки союзникам ценные сведения. Три… Два… Один.
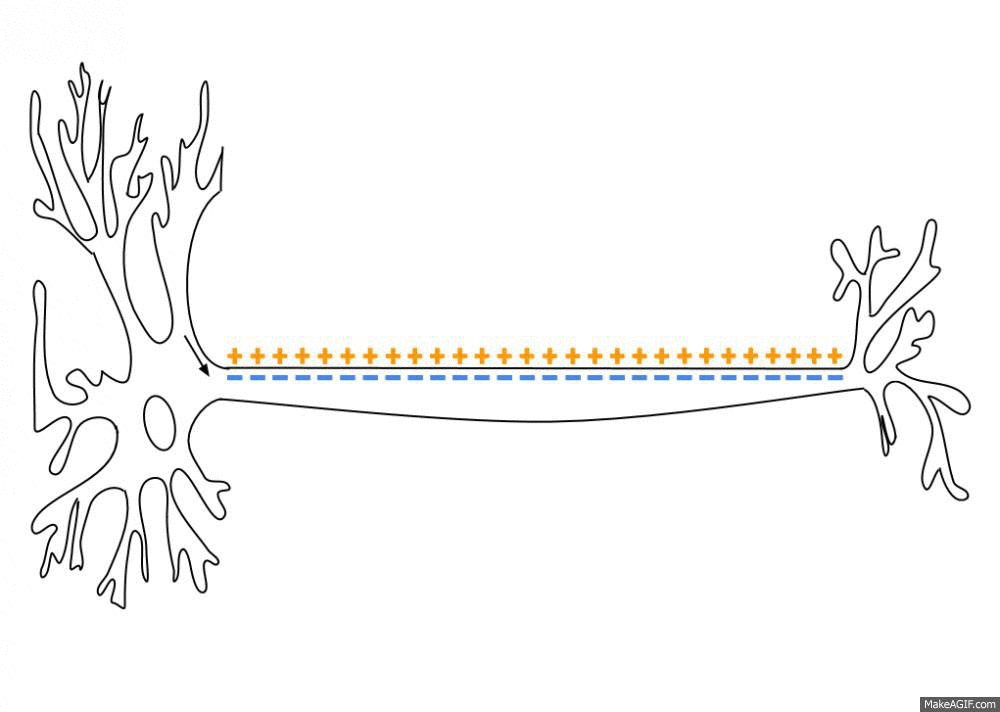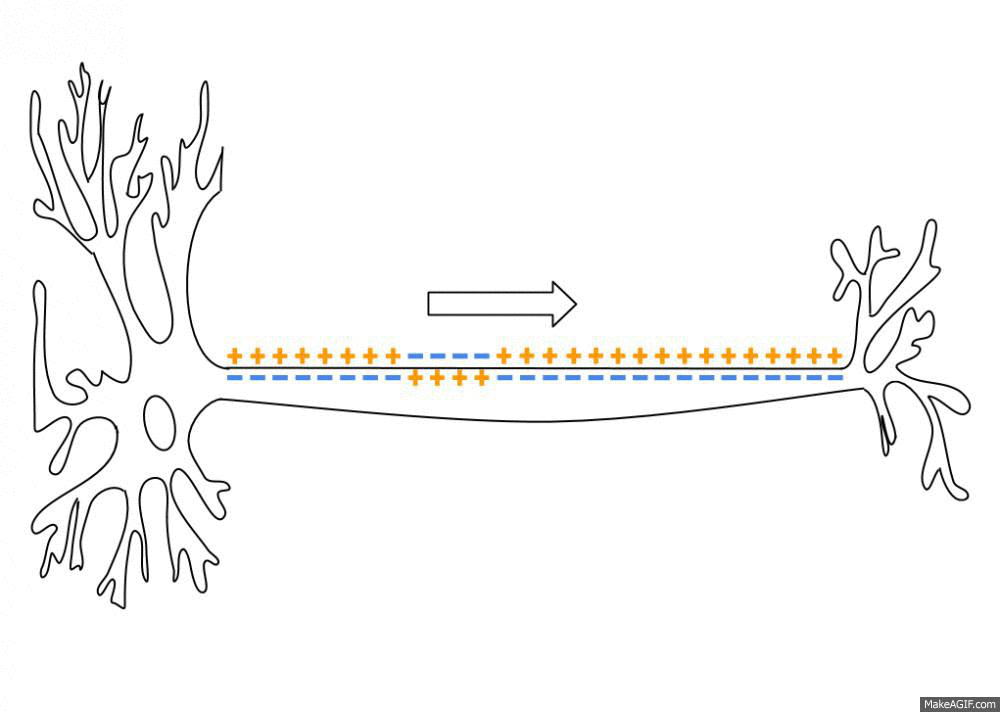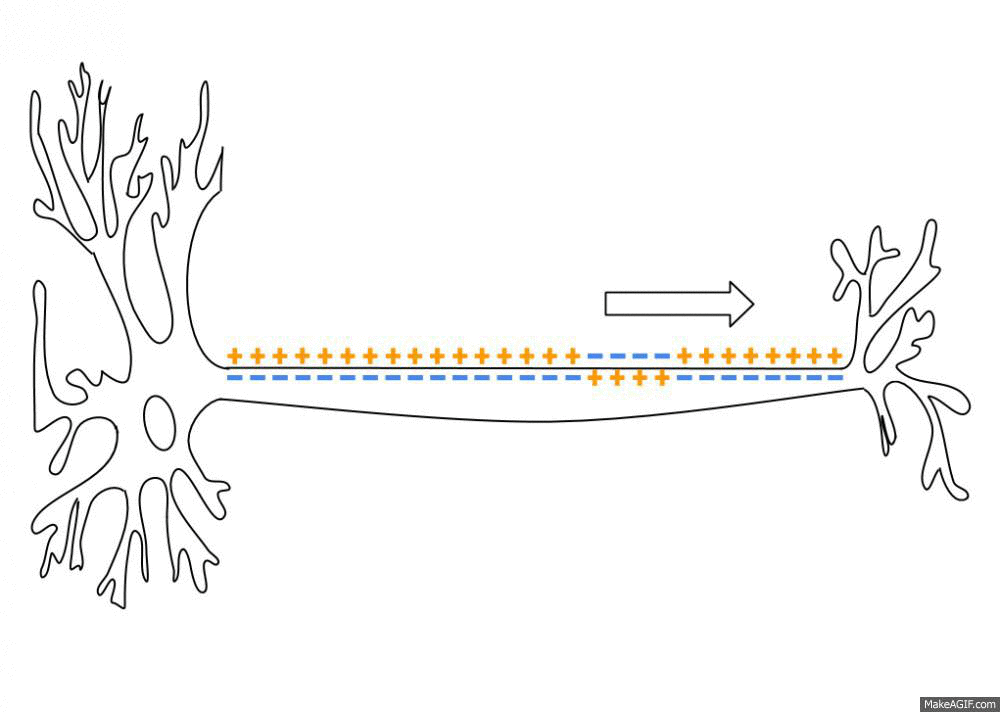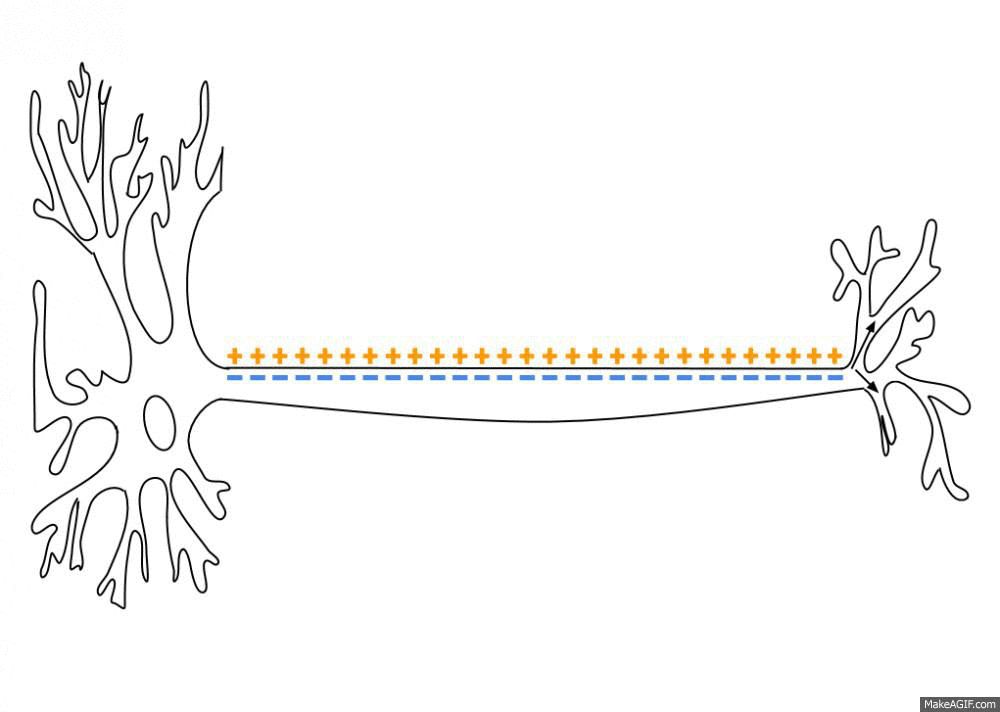
Распространение потенциала действия по аксону
Согласитесь, похоже на сценарий какого-то средневекового фильма? И всё это ежесекундно в огромных количествах происходит в каждом уголке нашего организма. Как вы, наверное, уже могли догадаться, речь идёт о потенциале действия (ПД) – другом двигателе, который обеспечивает нашу жизнедеятельность.
В прошлый раз мы говорили о потенциале покоя, который полностью противоположен нашему нынешнему «герою», но только в их постоянном чередовании может существовать чудо под названием «жизнь». Покой обеспечивает внутреннюю устойчивость и равновесие, которые можно регулировать и менять действием. Именно так «сотрудничают» потенциалы, то есть без одного невозможен и другой. Но давайте обо всём по порядку.
Мы с вами уже разобрались, какие бывают каналы и как они поддерживают внутреклеточный гомеостаз (постоянство среды). Калиевые каналы обладают одними «вратами», который в покое открыты, а натриевые – двумя, и те, которые располагаются более близко к наружной среде – в покое закрыты. Но вот как только клетка начинает возбуждаться и генерировать потенциал действия, всё меняется. Как? Двухфазно и циклически.
Дело в том, что потенциал действия тоже не возникает на «пустом» месте, и для того, чтобы он появился, нужен свой определённый по силе пороговый раздражитель. Здесь самое время сказать о том, что раздражители тоже бывают разными: одни подходящие (пороговые, их воздействие вызывает ПД), другие слабоваты (подпороговые, вызывают локальный ответ, но до возбуждения «не дотягивают»), а третьи – чрезмерные (надпороговые, могут и «убить»).

Итак, фаза 1 – деполяризация.
В покое мембрана поляризована, и мембранный потенциал (разница между зарядами внутри и снаружи) составляет у нервных клеток и волокон -70 мВ (для сравнения, у мышечных клеток он равен -90 мВ). Как только на клетку действует раздражитель (предположим, он по силе «идеален»), натрий, которого много вовне, устремляется внутрь через постепенно открывающиеся потенциал-зависимые ионные каналы и деполяризует мембрану до того самого порогового уровня (около -55 мВ), после которого лавинообразно и одномоментно распахиваются абсолютно все подобные каналы. Таким образом, мембрана резко меняет свой заряд, и там, где был (-), становится (+) – внутри, а там был (+), становится (-) — снаружи. Мембрана приобретает заряд около +40 мВ, и тот пик, пока она заряжена положительно (выше 0 мВ), называется овершутом (нет это не «очень крутой» шут, а такой термин).

С этого момента начинается фаза 2 – реполяризация.
Достигнув максимального пика, сила потенциала падает, и ионный ток начинает обратное движение (нужно же убрать «всё это безобразие» после «вечеринки»). При этом потенциал-зависимые натриевые каналы захлопываются, но открываются каналы для К+, который начинает активно выходить по градиенту концентрации (помним, что снаружи его меньше) и восстанавливать внутренний отрицательный заряд мембраны. Однако, натрия в клетке всё ещё много, и чтобы его ликвидировать, подключаются энергозатратные К/Na-насосы. В итоге все эти механизмы перевыполняют свою работу, и клетка слегка уходит в гиперполяризацию, приобретая даже более сильный отрицательный заряд, который ей в принципе нужен для покоя. Но потом всё быстро восстанавливается, и вот мембрана уже снова готова к приходу сокрушительного деполяризующего «цунами».
Конечно, стоит отметить, что вторая фаза потенциала действия, как правило, обусловлена не одним, а целым семейством взаимодействующих калиевых, кальциевых и других токов.
Какие же условия нужны для того, чтобы появился потенциал действия? Прежде всего, должно «прилететь» раздражение либо извне, либо благодаря блуждающим по мембране локальным потенциалам, которые, объединившись на аксональном холмике (его можно даже назвать «родильным домом»), переходят в реальный потенциал действия. С этого момента он становится нервным импульсом и передаётся по аксону, вызывая в синаптическом окончании выброс нейромедиаторов, которые устремляются к мембране другой клетки через синаптическую щель (узнаёте картину, описанную в начале статьи?)
Кроме того, то самое раздражение должно быть достаточным по силе, чтобы достичь порога возбуждения, достаточным по времени действия и скорости нарастания. Имеет значение даже его направление (если оно электрическое, то в зависимости от полюса направление будет либо в сторону деполяризации, либо в сторону гиперполяризации).
Так, стоп. А почему нас это волнует? Поясним: чем мембрана более деполяризована, тем клетка более активна и готова на ответ в виде потенциала действия. Например, если её начальный заряд не -70 мВ, а -60. Именно поэтому любое малейшее движение (зачёркнуто) раздражение, даже подпороговой силы, будет способно клетку возбудить и заставить её «вздрогнуть в конвульсии».
Когда же она находится в состоянии гиперполяризации, ей глубоко «всё равно», какими пороговыми раздражителями на неё воздействуют. Например тогда, когда мембранный потенциал составляет -100мВ, и до порога нужно преодолеть целых 45мВ. Она «не в духе», и только сила гораздо выше обычного для неё порога сможет «достать» её из этого состояния и помочь ответить потенциалом действия. Но уж как только она достигает этого порогового значения, то отвечает со всей той силой и «пылкостью», на которую способна (согласно закону «всё или ничего»). А измеряется уровень «пылкости» количеством имеющихся в мембране потенциал-зависимых Na-каналов, которые могут открыться одновременно.

Движение ионов через каналы в процессе распространения потенциала действия
Но и это ещё не всё. Оказывается, на разных стадиях развернувшегося потенциала действия клетка тоже поддаётся (или не поддаётся) влиянию. Если попытаться её тронуть на этапе быстрой деполяризации и сразу после пика, то ничего не произойдёт, потому что в первом случае все возможные Na-каналы уже будут открыты, а во втором – инактивированы (закрыты на обе «щеколды»). Этот период называется периодом абсолютной рефрактерности. Но если ещё немного подождать и попытаться пораздражать мембрану за овершутом, то мы увидим ответ, хотя он будет достаточно слабым – период относительной рефрактерности. Это потому, что некоторые натриевые каналы уже «оправятся» и наполовину откроются, готовясь к новой серии возбуждения. Сразу за ним идёт короткий период повышенной возбудимости, так как клетка ещё не «остыла» и готова откликнуться потенциалом на любой слабый сигнал.
Если же мы потерпим ещё немного, избежав период гиперполяризации, и дадим раздражение, то увидим, что мембрана снова гипервозбудима, потому что попадём в так называемый следовый потенциал.
Такие вот клеточные «американские горки».
Интересно, что амплитуда потенциала действия у каждой клетки постоянная и своя, правда, колеблется не так сильно и в основном составляет по абсолютной величине около 90-110 мВ. А вот длительность ПД у разных типов клеток различается и довольно значительно, вплоть до нескольких порядков.
Наиболее «быстрострельные» потенциалы действия у крупных миелинизированных аксонов двигательных нейронов (2-3 мс), а также скелетных мышечных волокон (3-4 мс). Оно и понятно – им важна скорость. Прежде всего, для выживания.
Гладкомышечные клетки генерируют потенциалы действия не всегда, а их продолжительность может составлять от 10 до 30 мс, как, например, в клетках желудочно-кишечного тракта. Здесь сила сокращения мышцы зависит от того, насколько часто возникают серийные потенциалы. Схема такая: чем чаще один ПД следует за другим, тем больше кальция поступает в клетки, и, соответственно, тем сильнее сокращается и тонизируется гладкая мышца.
У кардиомиоцитов (мышечных клеток сердца) длительность ПД больше на целых два порядка и составляет 300-400 мс, так как здесь во время второй фазы потенциала внутрь поступают ионы кальция, которые нужны для того, чтобы запустить сокращение. В итоге чем длительность ПД отдельной клетки выше, тем сильнее сокращение сердца.
Зачем нам всё это нужно знать? Всё достаточно просто. Потенциал действия и колебания потенциала покоя лежат в основе возбуждения и торможения у животных и человека. Главные характеристики ПД – амплитуда и длительность – могут быть как функциональными, так и носить патологический характер, что сразу даст нам понять о «сбоях в системе». А исследование суммарных потенциалов действия клеток и органов легло в основу таких незаменимых в современной врачебной практике диагностических методик, как электрокардиография, электроэнцефалография и электромиография.
Текст: Анна Хоружая
Предыдущие выпуски: